




Lupine Publishers Group
Lupine Publishers
ISSN: 2637-6652
Mini Review (ISSN: 2637-6652) 
The Perception of Fluctuations of Cell Velocity as a Possible Means of Overcoming Sedimentation by Motile Sea-Plankton Microorganisms Volume 2 - Issue 3
Received: December 21, 2018; Published: January 04, 2019
Corresponding author: Pundyak Oleh, Botanical Garden of Ukrainian National Forestry University, Ukraine.
DOI: 10.32474/MAOPS.2019.02.000139
Abstract
A model for overcoming the gravity by cells of wide range of motile sea-plankton microorganisms without gravity perception or antipredator behavior is proposed here. It is based on the assuming that the cells are able to detect thermal fluctuations of their velocities. In our model the cell locomotion force linearly depends on the cell velocity fluctuations causing especial type of gravikinesis. According to equations of the cell movement, sedimentation of the organisms may be considerably overcome.
Keywords:Microplankton; Picoplankton; Sedimentation; Gravikinesis
Introduction
The speed of some microorganisms depends on their orientation within the gravity field (excluding buoyancy of the whole cells). This phenomenon is called gravikinesis. Physiological mechanisms of gravikinesis have been well examined and described by Hemmersbach & Häder et al, [1] for example. It is expressed by the existence of certain cell receptors accepting gravistimulus and, through signaling chains, causing the change of cell motor work. The physical aspect of gravikinesis might be expressed by an anisotropic interaction of the microorganisms with the water medium. Recent calculations [2] supported our hypothesis that the physical aspect of gravikinesis can occur in bacterial and also in eukaryotic motile sea-picoplankton cells. According to our mentioned calculations the physical aspect of gravikinesis can be visible only in the case of high concentrations of bacteria without antipredator behavior (AB) equaling or bigger than 108 cells m-3 in consequence of friction anisotropy of co-existing microorganisms with high level of AB. Thus, a question can be raised: whether motile sea-plankton microorganisms in low concentrations can overcome gravity without any special cellular mechanism for gravity perception, as it has been shown for asymmetric microplankton [3-5] or for sea-picoplankton with antipredator behavior [2] ? We have tried to answer this question positively taking into account the existing of mechanosensitive channels, which can detect the changes of cell membrane tension linked with the changes of friction force during fluctuations of cell velocity [6]. Let us show the possible result when an averaged fluctuation force, which acts on the cells with ability to detect the thermal fluctuations of their velocities, is considerable in comparison with buoyance force. As the velocity fluctuations are very ephemeral (their duration for plankton organisms with dimensions up to 10-5 m does not exceed 10-5 s), they can be detected as the difference between the real cell velocities and averaged ones:

where 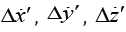 are the cell velocity fluctuations. Let us
consider such a velocity fluctuations detection in our model system.
For this goal let us transform equations (1), (2), (3) [2] having
considered equation (9) [2]. Thus, analogically to equation (11) [2]
we obtain:
are the cell velocity fluctuations. Let us
consider such a velocity fluctuations detection in our model system.
For this goal let us transform equations (1), (2), (3) [2] having
considered equation (9) [2]. Thus, analogically to equation (11) [2]
we obtain:

where β is the coefficient of the cell velocity fluctuations detection. If β =const , then it does not influence on the sedimentation rate. But it is natural to consider that the receptors of the velocity fluctuations detect only scalar velocity deviations. So, in our model |β| =constand when Δx′ change its sign, β does it too. There appear two variants: with positive and negative values of β ⋅Δx′ . The first corresponds to the inhibition of the cell motor as the respond on the deviations. The latter – to the stimulation. Thus, according to [2] during the averaging of equation (2) for Δx′ , averaged fluctuation force (let us call it diffusional force) may be not equal zero:

Thus, formula (18) [2] changes as

where dx’ and dy’,z’ are the dimensions of the particles along the axes X’ and Y’ or Z’ (of orthogonal coordinate system bounded with the axis (X’), along which the locomotion force always acts) correspondingly; Dx’ and Dy’,z’ are coefficients of diffusion along the axis X’ and Y’ or Z’ correspondingly. It is natural to assume that β ≥ hx’. Let us consider that
ξ ≈10.
Thus, according to all calculations done in [2] not only for picoplanktonic (as it was shown for the cells with antipredator behavior) but for a part of microplanktonic organisms (with the dimensions from 2·10-6 m up to 8·10-6 m) the diffusional force may have a considerable value in comparison with buoyant force. Thus, we can say, that picoplanktonic organisms and the part of microplanktonic ones with their dimensions up to 8·10-6 m, which have the cell velocity fluctuations receptors can considerably decrease sedimentation without any special receptors of gravity. We also can conclude that, though poorly studied generally and especially in the world of motile picoplankton and microplankton, the receptors of cell velocity fluctuations can also serve as an adaptation to overcome sedimentation. This finding may stimulate further research in this field.
References
- Hemmersbach R, Häder, DP (1999) Graviresponses of certain ciliates and flagellates. FASEB J 13: 69-75.
- Pundyak O (2017) Possible means of overcoming sedimentation by motile sea-picoplankton cells. Oceanologia 59 (2): 108-112.
- Hagen B, Kümmel F, Wittkowski R, Takagi D, Löwen H, et al. (2014) Gravitaxis of asymmetric self-propelled colloidal particles. Nat Comm 5(9): 1-7.
- Kessler JO (1985) Hydrodynamic focusing of motile algal cells. Nature 313 (1): 218-220.
- Roberts AM, Deacon FM (2002) Gravitaxis in motile micro-organisms: The role of fore-aft body asymmetry. J Fluid Mech 452(1): 405-423.
- Rasmussen T (2016) How do mechanosensitive channels sense membrane tension? Biochem Soc Trans 44(4): 1019-1025.


-

Mark E Smith
Bio chemistry
University of Texas Medical Branch, USA -

Lawrence A Presley
Department of Criminal Justice
Liberty University, USA -

Thomas W Miller
Department of Psychiatry
University of Kentucky, USA -

Gjumrakch Aliev
Department of Medicine
Gally International Biomedical Research & Consulting LLC, USA -

Christopher Bryant
Department of Urbanisation and Agricultural
Montreal university, USA -

Robert William Frare
Oral & Maxillofacial Pathology
New York University, USA -

Rudolph Modesto Navari
Gastroenterology and Hepatology
University of Alabama, UK -

Andrew Hague
Department of Medicine
Universities of Bradford, UK -

George Gregory Buttigieg
Maltese College of Obstetrics and Gynaecology, Europe -

Chen-Hsiung Yeh
Oncology
Circulogene Theranostics, England -
.png)
Emilio Bucio-Carrillo
Radiation Chemistry
National University of Mexico, USA -
.jpg)
Casey J Grenier
Analytical Chemistry
Wentworth Institute of Technology, USA -

Hany Atalah
Minimally Invasive Surgery
Mercer University school of Medicine, USA -

Abu-Hussein Muhamad
Pediatric Dentistry
University of Athens , Greece
