




Lupine Publishers Group
Lupine Publishers
ISSN: 2644-1217
Research ArticleOpen Access
Quantum Entanglement Entropy Causes Proliferating Metastases, Coincidences and Co-Occurrences of Cancer Volume 2 - Issue 2
Received: December 05, 2019; Published: January 23, 2020
*Corresponding author: Robert Skopec, Department of Biology-Psychology, Komensky University, Slovakia
Abstract
An important role in the processes of adaptation and masking in humans is playing also the immune system. The innate immune system functions as an interpreter of tissue damage and provides a first line of defense, also translates the information to other repair and defense systems in the body by stimulating angiogenesis, wound repair, and activating adaptive immunity. It is appropriate to consider autophagy a means for programmed cell survival balancing and counter-regulating apoptosis. Autophagy seems to have a dichotomous role in tumorigenesis and tumor progression.
Introduction
One prevalent description of translational medicine, first introduced by the Institute of Medicine’s Clinical Research Roundtable, highlights two roadblocks (i.e., distinct areas in need of improvement): the first translational block (T1) prevents basic research findings from being tested in a clinical setting; the second translational block (T2) prevents proven interventions from becoming standard practice. Bureaucracy is also a form of proliferating Social Metastasis.
Dichotomous correlations of cancer adaptation
Two other attributes play a similarly paradox role. The first involves major reprogramming of cellular energy metabolism in order to support continuous cell growth and proliferation replacing the metabolic program that operates in most normal tissues. The second involves active evasion by cancer cells from attack and elimination by immune cells. This capability highlights the dichotomous correlations of an immune system that both antagonizes and enhances tumor development and progression [1,2]. Evidence began to accumulate in the late 1990s confirming that the infiltration of neoplastic tissues by cells of the immune system serves counter-intuitively to promote tumor progression.
The bipolar nature of cancer: HYBRID, twofaced new main law of Nature
The quantum entanglement is a basis of twofaced reality in which we are living our lives. From this reality are outgoing also the science and healthcare too. Although metastasis is important for systemic correlations expansion (as in tumors), it is a highly dichotomous process, with millions of cells being required to disseminate to allow for the selection of cells-correlates aggressive enough to survive the metastatic cascade. To quantify the dynamics of metastasis of correlations development, we need look at the coincidences of metastases in terms of co-occurrence at every point of time [3,4] To quantify co-occurrence we can use the ϕ -correlation between dichotomous variables defined as:
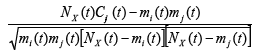 where C (t) ij is the
number of co-occurrence at time t.
where C (t) ij is the
number of co-occurrence at time t.
Then i and j represent particular site of metastasis, X represents
the primary correlations type. The pair-wise correlations
(coincidences) between metastasis network links for every primary
correlations types and lead to the correlation coefficient matrix.
The dichotomous correlations of the adaptation may be caused also
by the Quantum Entanglement Relative Entropy as a measure of
distinguishability between two quantum states in the same Hilbert
space. The relative entropy of two density matrices P0 and P1 is
defined as 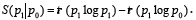 When P0 and P1 are
reduced density matrices on a spatial domain D for two states of
a quantum field theory (QFT), implies that
When P0 and P1 are
reduced density matrices on a spatial domain D for two states of
a quantum field theory (QFT), implies that 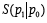 increases with
the size of D. Than
increases with
the size of D. Than 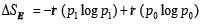 is the change in entanglement entropy across D as one goes between the states.
When the states under comparison are close, the positivity is
saturated to leading order:
is the change in entanglement entropy across D as one goes between the states.
When the states under comparison are close, the positivity is
saturated to leading order: 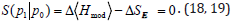
The problem of conventional adaptation may be given by a definition of static, deterministic world. The proliferative correlations of cancer lead to the resonances between the degrees of freedom. When we increase the value of energy, we increase the regions where randomness prevails. For some critical value of energy, chaos appears over time we observe the exponential divergence of neighboring trajectories. For fully developed chaos, the cloud of points generated by a trajectory leads to diffusion. Here we must as first formulate a new Main Natural Law: the HYBRID Quantum Entanglement Entropy (HQEE). Through above resonances the QEE is causing a metastasis of correlations, antagonistically intertwining (coincidences) all types of potentially conflicting interests in cancer.
The spatiotemporal entanglement evolution in free space
The many-boson interpretation may be applied to study of entangled photons as well, where the two-photon probability is used to obey the Wolf equations by Saleh, Teich, and Sergijenko (STS). Instead of treating the entanglement properties of the photons, and the optical propagation as two separate problems, with the STS equations, we can use now a single quantity-namely, the two-photon amplitude-to keep track of the spatiotemporal entanglement evolution in free space. This is analogous to the Wolf equations, which reformulate the laws of optics in terms of coherence propagation. We utilize the STS treatment of the two photons in study of various temporal effects. The Schrödinger picture would offer a more accessible interpretation of temporal entanglement propagation for studies of two-photon systems. For example, a four-wave mixing in a coherently prepared atomic gas, thus extending the STS model for use in many more topics in quantum optics in order to demonstrate the use and intuitiveness of the Schrödinger picture.
Based on this formalism, we propose the concept of quantum plasmatic temporal imaging, which uses dispersive elements and temporal phase modulators to manipulate the temporal entanglement properties of two photons. It is possible to convert positive-time correlation to negative-time correlation, or vice versa, using a plasmatic temporal imaging system. This conversion technique could be immensely useful for applications that require negative-time correlation, such as quantum-enhanced clock synchronization. Generating of negative-time correlation directly has some shortcomings compared with the conventional triedand- true schemes that generate positive-time correlation. This technique could allow more flexibility in choosing two-photon sources for quantum optics applications. We can consider two photons in two optical modes, such as two polarization, two propagation directions, or two waveguide modes [4-7].
The two-photon wave function is
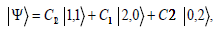
Where the constants 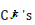 are overall amplitudes of the
quantum states,
are overall amplitudes of the
quantum states, 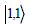 is the quantum state in which one photon is
in each mode,
is the quantum state in which one photon is
in each mode, 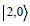 is the state in which both photons are in mode
1, and
is the state in which both photons are in mode
1, and 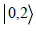 is the state in which both photons are in mode 2. The
positive-frequency forward-propagating component of the electric
field in each mode is given by
is the state in which both photons are in mode 2. The
positive-frequency forward-propagating component of the electric
field in each mode is given by
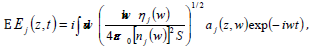
Where nj is the complex, frequency-dependent refractive index in mode j, nj is the real part of nj , S is an area of quantization in the x-y plane, and aj is the photon annihilation operator, related to the corresponding creator operator via the equal-space commutator
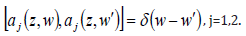
The physical significance of each amplitude  is that
its magnitude squared gives the probability density
is that
its magnitude squared gives the probability density 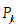 of
coincidentally measuring one photon in mode j at (z,t) and another
photon in mode k at (z′,t′ ),
of
coincidentally measuring one photon in mode j at (z,t) and another
photon in mode k at (z′,t′ ),
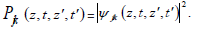
Plasmatic temporal entanglement is defined as irreducibility of 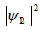 into a product of one-photon amplitudes in the form of a(t)b(
t′). This means that the probability of detecting a photon in mode 1
at time t is correlated to the probability of detecting a photon in mode
2 at t′. The most popular ways of generated entangled photons are
spontaneous parametric down-conversion and four-wave mixing,
where wave-mixing geometry and the spatiotemporal profile of the
pump-beam determine the initial
into a product of one-photon amplitudes in the form of a(t)b(
t′). This means that the probability of detecting a photon in mode 1
at time t is correlated to the probability of detecting a photon in mode
2 at t′. The most popular ways of generated entangled photons are
spontaneous parametric down-conversion and four-wave mixing,
where wave-mixing geometry and the spatiotemporal profile of the
pump-beam determine the initial 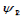 . The most interesting case
is when M=-1 and one of the photons is time reversed. If the two
photons are initially entangled with positive-time correlation, can be written as (0,t, z′,t′ )=a(t)b(t- t′), where b is assumed
to be much sharper than a. After photon 1 has passed through
the plasmatic temporal imaging system M=-1, (z,t, z′,t′ )=a(
td d t)b(t t t ) The photons hence become anticorrelated
in time. Since most conventional two-photon sources generate
positive-time correlation, but negative-time correlation is desirable
for many applications, one can use the temporal imaging system to
convert the former to the latter form of cancer.
. The most interesting case
is when M=-1 and one of the photons is time reversed. If the two
photons are initially entangled with positive-time correlation, can be written as (0,t, z′,t′ )=a(t)b(t- t′), where b is assumed
to be much sharper than a. After photon 1 has passed through
the plasmatic temporal imaging system M=-1, (z,t, z′,t′ )=a(
td d t)b(t t t ) The photons hence become anticorrelated
in time. Since most conventional two-photon sources generate
positive-time correlation, but negative-time correlation is desirable
for many applications, one can use the temporal imaging system to
convert the former to the latter form of cancer.
Besides the above application, one can also convert negativetime
correlation, which can be generated by ultrashort pulses
for improved efficiency, to positive correlation. Any desired
correlation can actually be imposed on already entangled photons,
by multiplying the original correlation with a factor of 1/M [8-12]. As group-velocity dispersion and temporal phase modulation play
analogous roles in the time domain to diffraction and lenses, which
can be use Fourier optics, temporal imaging, and quantum imaging
techniques to design more complex quantum plasmatic temporal
imaging systems. The quantum destructive interference via a
coupler is determined by the overlap of the two photons amplitude (0,t,0,t′) with its plasmatic mirror image.
The output amplitude is the destructive interference between the original amplitude and its replica but with the two photons exchanging their positions in time. In particular, for a 50%-50% coupler, T=R=1/2, complete destructive interference is produced if the two photons are temporarily indistinguishable. The introduction of variable distinguishability to photons, in order to produce varying degrees of destructive interference via a beam splitter to measure the two-coherence time, is the basic principle of the Hong-Ou-Mandel interferometer. As envisioned by Lukin and Imamoglu, the third-order nonlinear effects among two photons can become significant in a coherently prepared plasmatic atomic gas. The coupled-mode equations and then become nonlinear, where is the self-phase modulation coefficient and also cross-phase modulation coefficient. The advantage of the Schrödinger picture is most evident, whereas in the Heisenberg picture one needs to solve nonlinear coupled-mode operator equations, in the Schrödinger picture, one only needs to solve linear equations, which are similar to the configuration-space model applied to the quantum theory of solitons. The delta function δ (t − t′) couples the two subspaces, so entanglement can emerge from unentangled photons.
A soliton formed by two photons in orthogonal polarizations exerting cross-phase modulation on each other. Studies of two photons in the same mode under the self-phase modulation effect have been performed by entanglement, and a cross-phase modulation offers the distinct possibility of entangling two photons in different modes. Consider the case in which two polarizations have the same group-velocity dispersion, so that β2 = β2 = β21 , and there is one photon in each polarization. The evolution equation for
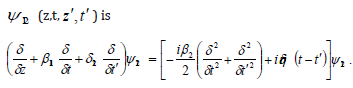
Defining time coordinates in a plasmatic moving frame of Cancer
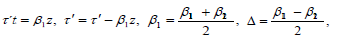
we obtain the following equation for (z,t, z′,t′ ):
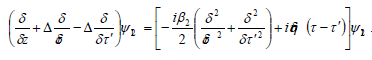
This equation is a simple linear Schrödinger equation,
describing a two-dimensional “wave function” (z,t, z′,t′ ) in a
moving frame subject to a δ ´potential. To solve for explicitly,
we define new time coordinates
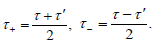
The cross-phase modulation effect only offers confinement of along the time difference ( − τ ) axis, but not the mean arrival
time ( + τ ) axis. The only bound-state solution of is
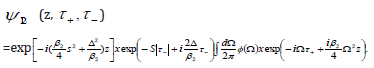
The delta potential enforces S to take on the value 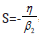 where
η and β2 must have opposite signs.
where
η and β2 must have opposite signs.
For the final solution in the cancer’s plasmatic moving
frame of τ and τ ′ is therefore
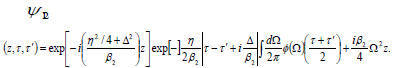
The two-photon coherence time of a vector soliton is fixed, but he average arrival time is still subject to dispersive spreading and becomes increasingly uncertain as they propagate. Hence a two-photon vector soliton generates temporal entanglement with positive-time correlation as it propagates. Similar to the idea of soliton momentum squeezing, one can also abadiatically change η,β2 along the propagation axis to control independently the two-photon coherence time. The center frequencies of the photons are shifted slightly, by an amount of + Δ/β2, to compensate for their group-velocity mismatch, so they can copropagate at average group velocity. This is commonly known as soliton trapping. If the nonlinearity has a finite band width, the potential becomes a finite-bandwidth function and multiple bound-state solutions can be obtained via conventional techniques of solving the linear the Schrödinger equation.
We have derived the general equations that govern the temporal evolution of two-photon probability amplitudes in different coupled optical modes. The formalism inspires the concept of quantum temporal imaging, which can manipulate the temporal entanglement of photons via conventional imaging techniques. The theory also offers an intuitive interpretation of two-photon entanglement evolution, as demonstrated by the exact solution of the two-photon vector soliton. To conclude, we expect the proposed formalism to be useful for many Plasmatic Moving Frames of Cancer, quantum signal processing and communication applications.
Why the universe includes the cancer at all
Don Lincoln, a senior physicist at Fermilab, does research using the Large Hadron Collider. He is the author of “The Large Hadron Collider: The Extraordinary Story of the Higgs Boson and Other Stuff That Will Blow Your Mind” and produces a series of science education videos. To the question: Why is there (in our Universe) something including cancer, rather than nothing? He is giving the simplest scientific answer: We shouldn’t exist at all. Give some scientists 65 pounds of rare isotope of germanium, cool it to temperatures cold enough to liquefy air, and place their equipment nearly a mile underground in an abandoned gold mine, and you’ll have the beginnings of an answer. Their project is called the Majorana Demonstrator and it is located at the Sanford Underground Research Facility, near Lead, South Dakota. To grasp why science has trouble explaining why matter exists-and to understand the scientific achievement of Majorana-we must first know a few simple things. First, our Universe is made exclusively of matter, all people, the Earth, even distant galaxies. All of it is matter.
Our best theory for explaining the behavior of the matter and energy of the Universe contradicts the realities that we observe in the Universe all around us. This theory, called the Standard Model, says that the matter of the Universe should be accompanied by an identical amount of antimatter, which, as its name suggests, is a substance antagonistic to matter. Combine equal amounts of matter and antimatter and it will convert into energy. And the street goes both ways: Enough energy can convert into matter and antimatter (since antimatter’s discovery in 1931). Modern cosmology says the Universe began in an unimaginable Big Bang-an explosion of energy. In this theory, equal amounts of matter and antimatter should have resulted in cancer. So how our Universe made exclusively of matter? However, we don’t know the process whereby the asymmetry in the laws of the Universe arose. One possible explanation revolves around a class of subatomic particles called leptons. The most wellknown of the leptons is the familiar electron, found around atoms. A less known lepton is called the neutrino. Neutrinos are emitted in a particular kind of nuclear radiation, called beta decay of cancer. It occurs when a neutron in an atom decays into a proton, an electron, and a neutrino. Neutrinos are fascinating particles. They interact extremely weakly, a steady barrage of neutrinos from the nuclear reactions in the Sun pass through the entire Earth essentially without interacting.
Still a mystery to scientists is whether there is a difference between neutrino matter and neutrino antimatter. While we know that both exists, we don’t know if they are different subatomic particles or if they are the same thing. We don’t know which kind of twins the neutrino matter/antimatter pair are. If neutrinos are their own antimatter particle, it would be an enormous clue in the mystery of missing antimatter and proliferation of matter in cancer. The way they do that is to look first for a very rare form of beta decay, called double beta decay. That’s when two neutrons in the nucleus of an atom simultaneously decay. If neutrinos are their own antiparticle, an even rarer thing can occur called HYBRID neutrino less double beta decay of cancer (the term: Robert Skopec). In this process, the neutrinos are absorbed before they get outside of the nucleus. In this case no neutrinos are emitted. The observation of a single, unambiguous neutrino less double beta decay would show that matter and antimatter neutrinos of cancer were the same.
If indeed neutrino less double beta decay exists, it’s very hard to detect and it’s important that scientists can discriminate between the many types of radioactive decay that mimic that of a neutrino. This requires the design and construction of very precise detectors. So that’s what the Majorana Demonstrator scientists achieved. Once and for all, it can answer the question of whether matter and antimatter neutrinos are HYBRID: the same or different. With that information in hand, it might be possible to understand why our Universe is made of matter leading to cancer too.
Majorana fermion change the world
Chinese scientists won a major victory, by proving that the Majorana fermion-a particle we’ve found tantalizing hints of for years-genuinely exists. This discovery has huge implications for quantum computing of cancer, and it might change the World. A Majorana fermion is weird even by the standards of quantum physics. The Majorana fermion doesn’t have a charge, which allows the mystery of cancer to be HYBRID: matter and anti-matter of cancer at the same time! The fact that it doesn’t have a charge, and also happens to be the exact reverse of itself to cancer at the same time. Quantum computers of cancer are like a huge pile of dimmer switches. You can set these dimmers much, much faster than you can flip on this light switches, because the dimmers are all wired to each other’s, immediately as tumors. These dimmers, e. i. quantum bits, are what’s called entangled in cancer. If you change one quantum bit, the others it’s entangled with change with it, even if they’re a million miles away from each other. That’s where Majorana fermions as metastasis come in due to their HYBRID: nocharge nature.
Qee as cancer
Only states that produce multiple informational offspringmultiple imprints on the environment-can be found out from the fragments of environment. The origin of the emergent classicality is then not just survival of the fittest states to deposit multiple records-copies of themselves-through environment. Proliferation of records allows information about system to be extracted from many fragments of the environment. Thus, environment acquires redundant records of the system [13-20].The mutual information is computed during the learning process learning. (See Eq in Figures below) Quantum mutual information (QMI) is defined in terms of the von Neumann entropy =as:. Unlike classical mutual information, the QMI between system A and B is not bounded by the entropy of either system. In the presence of the entanglement, the QMI can be as large as , which reflects the existence of quantum correlations beyond the classical one. Use of the naive Bayes classifier can be compactly represented as a Bayesian network with random variables corresponding to the class label , and the components of the input vector . The Bayesian network reveals the primary modelling assumption present in the naive Bayes classifier: the input attributes are independent given value of the class label. A naïve Bayes classifier requires learning values for the prior probability that the class label takes value, and the probability that input feature takes value given the value of a class label.
The feature selection filter in the perception can be used the naïve Bayesian classifier based on the empirical mutual information between the class variable and each attribute variable. The mutual information is computed during the learning process learning. (See Eq in Figures below). Kurt Gödel has pointed out, that predictions are like a perception of the objects of set theory. Prediction is a mode of mathematical intuition, which in sense of perception induces building up theories of the future. The given underlying mathematics is closely related to the abstract elements containd in our empirical ideas. The brain seems to have internal theories about what the world is like. Between brains theories is a internal perceptual rivalry in Darwinian sense. The World as a quantum system can be described due the polar decomposition, as a whole system consisting from two subsystems, which are mutually observing one another. During this observation the global workspace is processing reentry between internal representations and influence functional of the environment, between left and right hemisphere, etc. Some authors are proposing to consider a th of elements subset (X) taken from isolated neural system X, and its complement X - X (Edelman, 1998). Interactions between the subset and the rest of the system introduce statistical dependence between the two. This is measured by their mutual information MI(X, X - X) = H((X) + H(X - X)-H(X), which captures the extent to which the entropy of X is accounted for by the entropy of X - Xand vice versa (H indicates statistical entropy).
Decoherence as carcinogenesis
Another problem with the Darwinian adaptation is given by Quantum Darwinism and phenomenon of decoherence. It means that any coherent superposition of the system’s quantum states is continuously reduced to a mixture. A preferred basis called a “pointer basis” is singled out. An effective counterselection rule has emerged: the decoherence which prevents superpositions of the preferred basis from persisting by a lost of diagonal terms of the density matrix. Reduction to a mixture can’t be interpreted as adaptation, progress, development, forward evolution of the system. It is clearly only a counterselection in Western Powers.In this case the density matrix p(x,x’)of the particle in the position representations evolves according to the master equation (See Equation in Figures below) where H is the particle’s Hamiltonian, V(x) is potential, is the relaxation rate, is the Boltzmann constant, and T is the temperature of the field. This equation naturally separates into three distinct terms, each of them responsible for a different aspect of classical behavior: the first term, the von Neumann equation is derived from the Schrὅdinger equation. The second term causes dissipation: the loss of energy and decrease of the average momentum. The third term is responsible for the fluctuations or random kicks that lead to Brownian motion. They causing the environment-induced decoherence based counterselection.
Negative selection due to decoherence is the essence of environment-induced counterselection of Darwinian adaptation. Under scrutiny of the environment, only pointer states remain unchanged. Other states decohere into mixtures of stable pointer states that can persist, and, in this sense exist: they are counterselected.
Inhibitory-proliferative adaptation as cancer
In our model the values of and are transformed through a nonlinear activation function before they inhibit each other: (See Equation in Figures below). integration starts from , an input unit with mean activity , and independent white noise fluctuations of amplitude . These units also inhibit each other with a connection weight , denotes the decay rate of the accumulated activity with leak (entropy), means the number of alternatives. The inhibition parameter suppress the self-replicators, , - i.e. counterselect the.
Geniality and cancer : Neurobiological correlates of value have been described in orbitofrontal (conscience), cingulated cortex (critical intellectuals) and the basal ganglia, areas of the brain traditionally associated with reward-seeking behavior. Some neurons in orbitofrontal cortex represent value independently from evidence, choice and action. Anterior cingulate cortex is thought to represent negative (critical, non-linear) value. There is much evidence that a number of brain regions are sensitive to expected reward (or “utility”). The most well established are dopaminergic regions such as the striatum and midbrain structures. The common ratio pattern can be reconciled by the plausible assumption that people apply nonlinear decision weights to objective probabilities, so that the ratio is much smaller than. Neural responses to probabilities resembling the smoothly increasing function which typically fit behavior well. Paulus and Franck (2006) focused on between subjects measures and showed that activity in anterior cingulate correlated with degree of nonlinearity across subjects. We can make the assumption that neural activity is approximately a linear function of the behaviorally derived utility function. The GLM model separates the weighting function into two components: (1) component that is linear in and (2) the component that is the nonlinear deviation term (NDT) .
Specifically, we are looking for a prospect-theoretic expected value function that is nonlinear in ; that is . We assume the function is power function , where the value of is taken from the individual behavioral estimate, and , where the mean group =0.771 is used. If the expected utility (EU) null hypothesis is an accurate approximation of valuation of risky choices, there should be no reward-related brain regions that respond to the deviation term . If the nonlinear weighting hypothesis is an accurate approximation, there should be reward-related brain regions that respond equally strongly to the linear component and to the nonlinear component . We can test wheter cross-subject variation in the inflection of nonlinear weighting inferred from choices is consistent with cross-subject differences in neural activity. More highly nonlinear functions will be approximated by a combination of the linear term and the nonlinear term that puts more weight on the nonlinear term. A linear-weighting subject, will put no weight on nonlinear deviation. Denote the true weighting function for subject by, and the deviation from linear weighting by. A brain region that represents will be significantly correlated with both.
That is, the linear term and nonlinear deviation term with a higher weight on the nonlinear deviation term [13,14]. Brain regions that are significantly correlated with the nonlinear term include the anterior cingulated cortex (ACC), the striatum, motor cortex, and cerebellum. Our intuition is that brain activity during valuation of risks is more likely to correspond to cognitive components of prospect-masking, than to EU, and it will be easier to construct an adaptationist account of how evolution would have shaped brains to follow prospect-masking rather than EU.
The prospect-masking follows from psychophysics, while EU from normative logic. As a biological illustration of the subject with genial NDT we can propose Nobelist John Forbes Nash. He had new genial ideas, and later it has mentioned in his Nobel autobiography, because of “deviated somewhat from the line”. More exact definition of geniality is giving Robert Sternberg. On the top of that genius requires a combination of high motivation personality factors such as openness to experience, immense amounts of learning, at least ten years immersed in one’s discipline and environment that fits the potential genius like a glove. Many highly creative individuals lost a parent when they were young. This gives them an independent drive that might not have occurred otherwise.
The increased specialization required today for professional credentials makes the broad thinking of that characterizes geniuses harder to develop. I agree that the ritual culture of academia may also hamper genius. As philosopher of science Thomas Kuhn has pointed out, highly creative work (without precedent) does not fit existing formalistic academic paradigms tend to be dismissed (the counter-selection) [12,13]. Many great scientists have related how their most original ideas were repeatedly rejected by their peers. The most productive environment for the formation of new ideas, is one that encourages networks of minds operating in a non-market setting. For example, creative contributions that incrementally advance existing knowledge differ in their impact from those that redirect a field. The former are rewarded by a field’s referees and editors, the latter may be accepted only grudgingly, if at all, because they challenges the conventional wisdom. Inventive people also tend to be crowd-defiers. Creative people are thus intellectually combative. A working environment that encourages creativity must tolerate and even encourage such contrariness. It must also recognize that the more creative an idea is, the harder it will be to sell. Reviewers of grant proposals and journal articles must recognize that highly creative research may be less developed than that which only furthers established paradigms, and should make more allowances for originality.
Dichotomous correlations as cancer: One prevalent description of translational medicine, first introduced by the Institute of Medicine’s Clinical Research Roundtable, highlights two roadblocks (i.e., distinct areas in need of improvement): the first translational block (T1) prevents basic research findings from being tested in a clinical setting; the second translational block (T2) prevents proven interventions from becoming standard practice. An important role in the processes of adaptation and masking in humans is playing also the immune system. The innate immune system functions as an interpreter of tissue damage and provides a first line of defense, also translates the information to other repair and defense systems in the body by stimulating angiogenesis, wound repair, and activating adaptive immunity. It is appropriate to consider autophagy a means for programmed cell survival balancing and counter-regulating apoptosis.
Autophagy seems to have a dichotomous role in tumorigenesis and tumor progression. Two other attributes play a similarly paradox role. The first involves major reprogramming of cellular energy metabolism in order to support continuous cell growth and proliferation replacing the metabolic program that operates in most normal tissues. The second involves active evasion by cancer cells from attack and elimination by immune cells. This capability highlights the dichotomous correlations of an immune system that both antagonizes and enhances tumor development and progression. Evidence began to accumulate in the late 1990s confirming that the infiltration of neoplastic tissues by cells of the immune system serves counter-intuitively to promote tumor progression.
Twofaced new main law of nature as cancer: The quantum entanglement is a basis of twofaced reality in which we are living our lives. From this reality are outgoing also the science and healthcare too. Altough metastasis is important for systemic correlations expansion (as in tumors), it is a highly dichotomous process, with millions of cells being required to disseminate to allow for the selection of cells-correlates aggressive enough to survive the metastatic cascade. To quantify the dynamics of metastasis of correlations development, we need look at the incidence of metastases in terms of co-occurrence at every point of time. To quantify co-occurrence we can use the -correlation between dichotomous variables defined as: The dichotomous correlations of the adaptation may be caused also by the Quantum Entanglement Relative Entropy as a measure of distinguishability between two quantum states in the same Hilbert space. The relative entropy of two density matricesandis defined as . When and are reduced density matrices on a spatial domain D for two states of a quantum field theory (QFT), implies that increases with the size of D. Than is the change in entanglement entropy across D as one goes between the states.
When the states under comparison are close, the positivity is saturated to leading order: The problem of conventional adaptation may be given by a definition of static, deterministic world. The proliferative correlations lead to the resonances between the degrees of freedom. When we increase the value of energy, we increase the regions where randomness prevails. For some critical value of energy, chaos appears: over time we observe the exponential divergence of neighboring trajectories. For fully developed chaos, the cloud of points generated by a trajectory leads to diffusion. Here we must as first formulate a new Main Natural Law: the Quantum Entanglement Entropy (QEE) [15]. Through above resonances of the QEE is causing a metastasis of correlations, antagonistically intertwining all types of potentially conflicting interests [17].
Artificial carcinogenesis as cancer : For modeling cancer metastases spread there is several new methodologies. The nodes of a cancer metastasis network represents the distant sites where metastases could arise for a given tumor type. The size of each node represents its conditional incidence or hazard. The incidence hazard function is where is the number of diagnoses of metastasis met at time and is the number of patients remaining at time for primary tumor type [19]. The cummulative hazard from and met pair is to quantify the dynamics of metastasis development, we need the incidence of metastases in terms of co-occurence at every point of time. This allows to istablish links between the primary tumor and metastasis sites, as between different metastasis sites for multiple cases. With the fractional metod as a baseline for comparison was developed an algorithm for predicting future sites of metastases using cancer metastasis networks. These networks are entities on which the metastatic disease of individual patients evolve, and are able to incorporate temporal dynamics, and subtle relational properties.
Between anti-inflammatory receptors on macrophages of particular interest is CD200, whose by the endogenous ligand CD200L sends stop signal to macrophages suppressing production of proinflammatory mediators. CD200L expressed on activated immune cells provides a mechanism to dampen macrophage activation after initiation of the inflammatory response. Another receptor involved in negative regulation of inflammation is the recepteur d’origine nantis (RON), or stem cell-derived tyrosine kinase (STK) receptor in the mouse. The ligand for this receptor is macrophage-stimulating protein (MSP), a serum protein generated during the coagulation cascade. Even TLRs have the capacity of inducing expression of anti-inflammatory mediators, like IL- 10. The signaling pathway used by TLRs to activate expression of pro- and anti-inflammatory cytokines diverges at the level of the TRAF3 and TRAF6 proteins, because TRAF3 is critical for induction of IL-10 expression. In its absence, expression of the TRAF6-dependent proinflammatory cytokines IL-6 and IL-12 is dramatically upregulated.
Recent experiments have shown that nutrient starvation may induce intense autophagy which is causing cancer cells to shrink and adopt a state of reversible dormancy. These cells may exit from this state and resume active growth and proliferation after changes in tissue microenvironment (access to more nutrients). Each type of disseminated cancer cell needs to develop its own set of ad hoc solutions to the problem of thriving in the microenvironment of foreign tissue.
Executioner protein causes cancer cells to self-destruct : Researcher have discovered a way to use the executioner protein BAX to induce apoptosis in cancer cells while leaving healthy cells intact. The treatment has so far been applied only to acute myeloid leukemia (AML) cells but may have broader uses. NIH Albert Einstein College of Medicine ssresearchers have induced cancer cells to commit suicide with a new compound that leaves healthy cells untouched. They deployed their novel treatment approach against AML cells, which kill more than 10,000 Americans, and makes up about one-third of all new cases of leukemia, each year. Patients survive AML at a rate of only about 30%, making effective new treatments a hot commodity. Altough the team has only tested the treatment on AML, it could have the potential to succesfully attack other varieties of cancer cells. Researchers are hopeful that the targeted compounds they’re developing will prove more effective than current anti-cancer therapies by directly causing cancer cells to self-destruct, associate professor of medicine and biochemistry and senior author Evripidis Gavathiotis said in a press release. Ideally, this compounds would be combined with other treatments to kill cancer cells faster and more efficiently-and with fewer adverse effects, which are an all-too-common problem with standard chemotherapies.
The new compound fights cancer by triggering apoptosis: a natural process the body uses to get rid of maffunctioning and unwanted cells. Apoptosis also takes place during embryonic development: trimming excess tissue from the growing embryo. While certain existing chemotherapy drugs induce apoptosis indirectly by damaging the DNA in cancer cells, this new treatment directly triggers the process intentionally by activating BAX, the executioner protein. Pro-apoptotic proteins activate BAX in cells. Once BAX molecules go to work, they find the mitochondria of target cells and drill lethal holes into them, scuttling their ability to produce energy. Cancer cells resist BAX and this process by producing large quantities of anti-apoptotic proteins that suppress BAX and even the proteins that activate it. Theprocess discovered by the team of professor E. Gavathiotis wakes BAX again and sends it back to work against cancer cells. Their novel compounds revives suppressed BAX molecules in cancer cells by binding with high affinity to BAX’s activation site. BAX can then swing into action, killing cancer cells while leaving healthy cells unscathed.
In 2008, Dr. Gavathiotis was part of the team that first described the BAX’s activation site’s shape and structure. He has been searching for small molecules to activate BAX and produce sufficient activity to overpower the natural resistance cancer cells mount to apoptosis. His team screened more than one million compounds and narrowed the field to 500, many of them synthesized by the team, and then evaluated them. These results reveal the outcome of that search. BTSA1 (BAX Trigger Site Activator 1), was the best compound against several diffrent human AML cell lines, including those found in high-risk AML patients. BTSA1 was also able to induce apoptosis in AML cells without affecting healthy stem cells. In AML mice treated with the compound, there was a significantly longer survival rate: 43% of to control group was alive and AMLfree after 60 days. The BTSA1-treated mice also exhibited no signs of toxicity.BTSA1 activates BAX and causes apoptosis in AML cells while sparing healthy cells and tissues-probably because the cancer cells are primed for apoptosis. Next the team plans to test BTAS1 on other types of cancer using animal models Gavathiotis et al. [2].
Artificial metastatic colonization as Cancer : Macroscopic metastases may erupt decades after a primary tumor was surgically removed, or pharmacologically destroyed. These metastatic growths evidently reflect dormant micrometastases that have solved, after much trial and error, the complex problem of tissue colonization. This may support our hypothesis that tumor like an organ can be seen as a UTM programmed to make decisions outgoing from the Hayflick limit and trying to solve the Entscheidungsproblem. These adaptations require hundreds of distinct colonization programs, each dictated by the type of disseminating cancer cells and the nature of the tissue microenvironment. The colonization we can model with vectors. Let be a vector of observed variables, be a vector of latent variables, and be the model parameters. Lwt be a vector of all variables in the model. If were completly observed we could apply standard maximum likelyhood estimation to obtain:
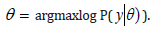
The is unobserved, becomes a random variable and there must be apply the Expectation Maximization (EM) algorithm. For more complex models, the free energy approach leads to more flexible model fitting. The standard EM is equivalent to performing coordinate ascent on the energy function
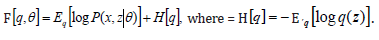
Beyond the timing we must know where cancer cells achieve the ability to colonize foreign tissue as macroscopic tumors. It may be during primary tumor formation when these cells enter into the circulation and are fortuitously endowed with capability to colonize distant tissues. The ability to colonize may also develop in response to the selective pressure [2] on disseminated cancer cells to adapt to growth in foreign tissue microenvironment. Tissue–specific colonization programs are evident in cells within primary tumor, may originate from emigrants returned home. Bureaucracy is also a form of Social Metastasis. The phenotypes and underlying gene expression programs of cancer cells within primary tumors may be modified by reverse migration of their distant metastatic progeny. In this self-seeding process the supportive stroma (arising in a primary tumor) contributing to acquisition of its malignant traits make possible reseeding and colonization by circulating cancer cells from metastatic lesions. The regulatory programs enabling metastatic colonization emerging in an important agenda for future research. Above metioned multitude of colonization programs are unlikely to depend only on cell-autonomous processes. The process of colonization probably encompass a large number of cell-biological programs, which are nonlinear and diverse [21]. The numerous signaling molecules affecting cancer cells as nodes and branches of the elaborate integrated circuits which are reprogrammed derivatives of the circuits operating in normal cells.
Bacterial starvation and general stress responses also promote mutagenesis during stress.Include the stringent and competence starvation-stress responses in Bacillus subtilis, and RpoE membrane-protein stress response in E.coli. These examples illustrate the multiple evolutions of mechanisms that couple genomic instability pathways with stress responses and stress. The importance of all of these is that genetic diversity is generated preferentially when cells are maladapted to their environment: when stressed.
Conclusion
An important role in the processes of adaptation and masking in humans is playing also the immune system. The innate immune system functions as an interpreter of tissue damage and provides a first line of defense, also translates the information to other repair and defense systems in the body by stimulating angiogenesis, wound repair, and activating adaptive immunity. It is appropriate to consider autophagy a means for programmed cell survival balancing and counter-regulating apoptosis. Autophagy seems to have a dichotomous role in tumorigenesis and tumor progression.
Conflict of Interests Disclosure
The author declares no conflict of interests
Tables
The mutual information

The mutual information 2

The density matrix p(x,x’)of the particle in the position representations evolves according to the master equation
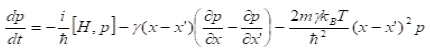
In our model the values of and are transformed through a nonlinear activation function before they inhibit each other:
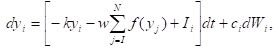
To quantify co-occurrence we can use the -correlation between dichotomous variables defined as:
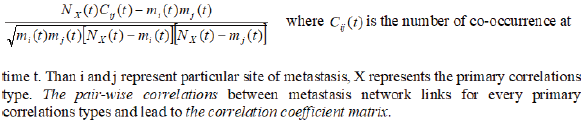
References
- Allen G, Chan T (2017) Artificial Intelligence and National Security. A study on behalf of Dr. Jason Matheny, Director of the US. Intelligence Advanced Research Projects Activity (IARPA); The Harvard University Belfer Center, USA, pp. 130-132.
- Gavathiotis E (2017) NIH Press Release , Eurekalert. Nature.
- Gong Bo Zhao (2017) Dynamical dark energy in light of the latest observations. Nature astronomy 1: 627-632.
- Liebig M (2003) Executive Intelligence Review Special Report. Washington DC, USA, p: 7.
- Rampino MR (2017) Cataclysms: A New Geology for the 21st. Century. New York University,USA.
- Skopec R I (2017) Coding By Quantum Entanglement Entropy. American Research Journal of Bioscience 2(3): 1-7.
- Skopec R (2017) De Programming Colorectal Cancer by Increase in Cholesterol Level. International Journal of Cancer Research & Therapy 2 (1): 1-4.
- Skopec R (2017) Agriculture As Producer of World Cancer Epidemic. MOJ Biology and Medicine 1(6): 00034.
- Vikman A (2005) Phys Rev D 71: 023515.
- Tennenbaum J (1988) Electromagnetic-Effect Weapons: The Technology and the Strategic Implications. Executive Intelligence Review, Wiesbaden, Federal Republic of Germany.
- Zhirinovsky A (2003) Speech at reception, Embassy of Iraq to Russia, Personal communications of diplomats in Moscow, Russia.
- Skopec R (2019) Quantum Resourrection: Quantum Algorithm With Complex Conjugation Reverses Phases Of The Wave Function Components. Journal of Neuroscience and Neurological Surgery.
- Skopec R (2018) Evolution Continues with Quantum Biology and Artificial Intelligence. ARC Journal of Immunology and Vaccines 3(2): 15-23.
- Skopec R (2019) Naphazoline nitrate treat the frey effect of microwave and other sonic weapon’s damages in human’s internal, organs. Virology: Research & Reviews 2(1):1-5.
- Skopec R (2019) Negative Health Effects of the International Space Station. Stem Cell Research International 3(2): 1-6.
- Skopec R (2019) Fifth Dark Force Completely Change Our Understanding of the Universe. Research Journal of Nanoscience and Engineering 3(2): 22-29.
- Skopec R (2019) Darwin’s Theorem Revised: Survival of the Careerist. Advancements in Cardiovascular Research 1(5): 89-93.
- Skopec R (2019) New Psychological Weapons Make Targets Hallucinate. Journal Of Neuropsychiatry And Neurodisorders 1(1): 1-6.
- Skopec R (2019) The Transfiguration with Self-Phase Modulation Effect of Entanglement in a Plasmatic Moving Frame. Current Trends in Biotechnology and Biochemistry.
- Skopec R (2019) Quantum Entanglement Entropy as Karma Lead to New Scientific Revolution. Journal of Engineering and Applied Sciences Technology 1: 1.
- Skopec R (2019) Quantum Entanglement Entropy Produces Energy by Info-Entropy Fields Forces. Modern Approaches on Material Science.


-

Mark E Smith
Bio chemistry
University of Texas Medical Branch, USA -

Lawrence A Presley
Department of Criminal Justice
Liberty University, USA -

Thomas W Miller
Department of Psychiatry
University of Kentucky, USA -

Gjumrakch Aliev
Department of Medicine
Gally International Biomedical Research & Consulting LLC, USA -

Christopher Bryant
Department of Urbanisation and Agricultural
Montreal university, USA -

Robert William Frare
Oral & Maxillofacial Pathology
New York University, USA -

Rudolph Modesto Navari
Gastroenterology and Hepatology
University of Alabama, UK -

Andrew Hague
Department of Medicine
Universities of Bradford, UK -

George Gregory Buttigieg
Maltese College of Obstetrics and Gynaecology, Europe -

Chen-Hsiung Yeh
Oncology
Circulogene Theranostics, England -
.png)
Emilio Bucio-Carrillo
Radiation Chemistry
National University of Mexico, USA -
.jpg)
Casey J Grenier
Analytical Chemistry
Wentworth Institute of Technology, USA -

Hany Atalah
Minimally Invasive Surgery
Mercer University school of Medicine, USA -

Abu-Hussein Muhamad
Pediatric Dentistry
University of Athens , Greece
