




Lupine Publishers Group
Lupine Publishers
ISSN: 2641-1768
Research ArticleOpen Access
Losses Loom Larger Than Gains: Neural Correlates Of Behavioral Gain-Loss Asymmetry Volume 5 - Issue 4
Received:August 02, 2021 Published:August 11, 2021
Corresponding author:Harold Miller, Department of Psychology, Brigham Young University, Provo, UT 84602, USA
DOI: 10.32474/SJPBS.2021.05.000217
Abstract
We measured the asymmetry of monetary gains and losses behaviorally using a computer game during which we simultaneously recorded participants’ electroencephalographic activity. The behavioral measures demonstrated that, on average, losses loomed larger than gains did (3.2:1). Event-related potentials, particularly the P300, had significantly greater mean amplitudes when losses occurred compared to gains (1.99:1) and correlated directly with the behavioral asymmetry we measured in the game (r = 0.85).
Keywords:Prospect Theory; Computer Game; Conjoint Interdependent Concurrent VI VI Schedule; Gain (Reinforcer); Loss (Punisher); Gain/Loss Asymmetry; P300 Wave; Event-Related Potential (ERP); EMOTIV EPOC+®; EEG; Generalized Matching Law
Introduction
Thomas Hobbes (1588-1679) proposed that voluntary behavior is governed by the principle of hedonism, that is, an individual’s sole intrinsic good is the overall pursuit of pleasure. A hedonist strives to maximize net pleasure by minimizing pain. Utility theory, which is a cornerstone of the rational perspective of economics, is rooted in the hedonist principle. However, the psychology of Homo economicus-a rational and self-interested individual with relatively stable preferences-has been challenged by numerous psychologists and behavioral economists. The purpose of our research was to explore the effects of small monetary gains and losses on choice behavior using a computerized game and to determine gain/loss ratio differences using both behavioral and electroencephalographic (EEG) measures.
A prominent example of gain-loss asymmetry is that losses loom larger than gains [1-4], meaning that the aversion to a loss of a certain magnitude is greater than the attraction to a gain of the same absolute magnitude. Such asymmetry is an indication that humans are sometimes biased in their decision making. Accordinng to Kahneman and Tversky [9], “The asymmetry of pain and pleasure is the ultimate justification of loss aversion in choice” [p. 157]. Kahneman, Knetsch, and Thaler [8] reported that “The existing evidence suggests that the ratio of the slopes of the value function in two domains, for small or moderate gains or losses of money, is about 2:1” [p. 199].
Rasmussen and Newland [14] reported a behavior-analytic experiment in which participants played a customized computer game that sometimes-produced reinforcers (gains) and sometimes punishers (losses) in the form of points exchangeable for money or, conversely, the loss of points and money. Their design included two alternating conditions. In one condition, a pair of reinforcement schedules were concurrently available. The ratio of reinforcer frequencies was adjusted systematically. The other condition consisted of the same pairs of reinforcement schedules, but a schedule of punishment was overlaid onto one of the reinforcement schedules. The authors reasoned that, in this way, they could measure the effect of punishers as the difference in the response ratios when punishers were delivered on one of the alternatives versus when punishers were absent from both alternatives. They concluded that the mean asymmetry ratio was approximately 3:1 (loss:gain) on average.
Kahneman [7] asserted that “the brain’s response to variations of probabilities is strikingly similar to the decision weights estimated from choices” [p. 315]. The electroencephalograph (EEG) may be used to record scalp visual-evoked potentials (VEPs), including event-related potentials (ERPs) [3,11]. Sokol-Hessner and Rutledge [15] reported that “Research on the neuroscientific basis of loss aversion has identified several critical neural components, suggesting a model of loss aversion in the human brain and providing links to the neuroscience of affect” [p. 315]. They reviewed several loss-aversion studies of young adults and adults and found that the means of loss aversion (computationally formalized as a multiplicative weight on losses relative to gains) in their model were between 1.3 and 2.5.
Kahneman and Tversky’s research method was cognitive. Rasmussen and Newland’s method was behavioral as was ours. However, we added an electrophysiological measure of the asymmetry of gains and losses. In the electrophysiological component of our study, the amplitudes of P300 waves elicited by gains and losses while playing the video game were measured in real time and converted to a gain/loss ratio. We hypothesized that the amplitudes of P300 waves recorded during a gain/loss behavioral procedure, when expressed as gain:loss ratios, would be directly related to their behavioral counterparts.
Method
Participants
The participants were 16 male undergraduate students enrolled at Brigham Young University (BYU), Provo, UT, USA 84602. They were recruited through an online recruitment platform following approval of our research protocol by the BYU Institutional Review Board.
Materials and procedures
We developed a computer game to produce behavioral data. Participants played the game in an experimental room, 9 ft by 9 ft, containing a table and chair. The table held a Dell® desktop computer (the game computer) equipped with a 17-in monitor and a mouse. The room was windowless and artificially illuminated. The computer had an Ethernet connection to a separate, identical computer that was in an adjacent room and that hosted the Emotive EPOC® Brainwear® software for recording the EEG and to monitor its functioning. The Emotive EPOC® device was placed on the participant’s head and contained 14 scalp electrodes. Written informed consent was obtained from all participants prior to the first experimental session. The participant was seated in front of the computer monitor and asked to read the instructions (written in English) for the game that appeared there. He was invited to play the video game in a series of 36-min sessions in which he could earn points on the screen. The net earnings were paid to the participant at the end of each session. In addition, the participants received a $50 bonus at the completion of the study. There were seven sessions. EEG recording was continuous during each session.
The SubSearch Game
Participants played SubSearch using the computer mouse to guide an underwater submarine and to retrieve as many yellow objects as possible before reaching the sea floor. When the cursor rested on the submarine, moving it moved the submarine. If the submarine was placed over a yellow object, clicking the mouse retrieved the object. Underwater barriers complicated the submarine’s movement between objects. Once the submarine descended to the sea floor, it was returned to the surface for a new descent, this time with more frequent barriers. Thus, the game became progressively more difficult as it continued. Only one panel was operative at a time. The other panel was darkened, and motion was paused. The game was played in two different vertical panels separated by a vertical line. Each panel was associated with its own interdependent concurrent variable-interval (inter conc VI VI) schedules of reinforcement. There was also a conjoint VI schedule of punishment during certain conditions on the left side of the screen. Unlike the traditional conc VI VI schedule in which the two schedules are independent of each other, the interdependent version assigned a reinforcer according to a preset probability generator. If, for example, the generator was set to assign twice as many reinforcers to the left panel than to the right panel (pL = 0.67), and the next reinforcer was assigned to the right panel, then it would be necessary for the participant to produce that reinforcer before the next one would be assigned. Thus, the interdependent schedule reduced the likelihood of extreme position (left or right) biases and assured that the scheduled proportion of reinforcers) between the two panels remained close to the proportion of those that were delivered.
After the participant clicked the “Start-OK” message on the screen, a 36-min session commenced. The game allowed the participant to move the cursor from one panel to the other. However, each switch produced a changeover delay of 2 s. During this interval, no reinforcers or punishers were delivered. Gains and losses were signaled by separate on-screen messages, each accompanied by a distinctive sound. For 0.5 s prior to the on-screen signal of a gain or of a loss, a fixation signal (a white + sign) was presented on the screen and followed in the same location by a message indicating either “Collect a coin to continue” for gains or “Insert a coin to continue” for losses. The gain-message appeared for 1 sec. Then the tab located at the bottom of the screen between the two counters began to blink. The game resumed after the participant clicked on the tab. Counters on each side of the tab displayed the net points for the respective side of the screen.
Each click during a session was coded, time stamped, and saved to an external MySQL database. The summary statistics included the total time spent responding in each panel, the total number of clicks that occurred in each panel, the total numbers of reinforcers and punishers that occurred in each panel, and the total number of changeovers. Each session consisted of a fixed sequence of six 6-min conditions (conditions 1-6). Three of them (1, 3, and 5) contained conc VI VI schedules of reinforcement only and three (2, 4, and 6) contained conc VI VI schedules of reinforcement and a conjoint VI schedule of punishment on the left side of the screen. Table 1 summarizes the scheduled frequencies of reinforcers and punishers in each condition. Condition 1 featured a conc VI60-s VI20-s schedule, meaning that 25% of the total reinforcers were allocated to the left panel and 75% to the right panel. There was no schedule of punishment. Condition 2 featured the same conc VI60-s VI-20 schedule of reinforcement plus a VI60-s schedule of punishments. In other words, 100% of the punishers were allocated to the left panel and no punishers to the right panel. The other four conditions featured different reinforcer ratios. Each unpunished condition was followed by a similar condition that included punishers only in the left panel under the same schedule as the reinforcers that were delivered in that panel. Each condition was accompanied by a different background color in each panels, for a total of six different colors. It should be noted that the values of the VI schedules in each concurrent pair of reinforcement schedules were selected to produce the same overall rate of reinforcement despite the difference in their ratios (1:3 in conditions 1 and 2, 1:1 in conditions 3 and 4, and 3:1 in conditions 5 and 6). The ratio of reinforcers to punishers was always 1:1.
Electrocortical activity
The head-mounted instrument was a wireless Bluetooth® Smart device (2.4GHz band) with 14 electrodes that transmitted at a sample rate of 128 Hz. It provided access to raw, densearray, high-quality EEG data with software subscription (EMOTIV Brainware®. The resolution was 14 bits with 1 LSB = 0.51 μV). The bandwidth was 0.2 -43Hz with digital notch filters at 50 Hz and 60 Hz. It included a digital 5th-order Sinc filter and a dynamic range (input referred) of 8400μV. It was AC coupled and powered by a lithium polymer battery (480 mV). The device sent the EEG data via a Bluetooth® connection to the computer to be recorded. In the game computer, certain in-game events, such as displaying a gain or a loss message on the monitor, triggered a signal to the second computer, it also compiled the data, temporally aligning the EEG data with the 8-bit codes received from the game and saved them to the hard disk. Because of the limitations of Bluetooth® range, both computers were in the same room, but the interface and the monitor for the second computer was in an adjacent room. The final output was a large csv file that contained a time-step column, the 14 electrode channels, and markers for each SubSearch on-screen message.
Event related potential analysis
VEPs are electrical potentials initiated by brief visual stimuli and are recorded from the scalp overlying the visual cortex. Amplitude (measured in μV) is defined as the difference between the mean pre-stimulus baseline voltage and specific voltages (positive and negative) measured within a time window. Latency (measured in ms) was defined as the time from stimulus onset to the point at which amplitude was measured within the window [11]. The ERP contains distinct waveforms that may be correlated with specific cognitive activities [2]. The labels N50, P100, N100, P200, N200, and P300 are commonly used, where P and N indicate positive or negative deflections, respectively, and the number indicates an ordinal position in the waveform. It should be noted that the P200, N200, and P300 are specifically ERPs; however, we used the term ERP to refer to all of the VEP components. Gehring and Willoughby [5] recorded brain activity coincident with monetary gains and losses and concluded that the amplitude was greater for losses than for gains. The ERP data were imported using EEGLab® with the ERPLab® add-on. EEGLab® is an interactive Matlab® toolbox for processing continuous and event-related EEG, magnetoencephalographic, and other electrophysiological data to produce independent component analysis, time/frequency analysis, artifact rejection, event-related statistics, and several modes of visualization of the averaged and single-trial data. A 1Hz high-pass filter, followed by a 50 Hz low-pass filter, was applied to the in-session recordings. Epochs were created for each gain or loss in the SubSearch game and ranged from 1,000 ms before the message appeared to 2,000 ms after it disappeared. Any epoch that contained an amplitude exceeding 150 mV was rejected. The epochs were averaged for gains and losses separately, resulting in a pair of summative waves (gain and loss) for each participant in each session. Then grand averages were created. The P300 component of the VEP was the focus of our analysis. The P300 wave was measured as the maximum positive deflection occurring between 250 msec and 500 msec following the presentation of brief visual stimulus. Yeung and Sanfey [17] found that, in studies of choice, the P300 can be influenced by several factors, including the magnitude of the chosen option, the valence and magnitude of the alternative option, and the relative value of the alternative outcome in comparison with the chosen outcome.
Table 1: Programmed Numbers of Reinforcers Per Minute and Punishers Per Minute in Each Condition of the SubSearch Game.

The data analysis consisted of signal filtering, amplifying, and averaging the EEG during the 1-s epoch immediately prior to the onset of a message on the monitor screen and during the 2-s epoch following the offset of the message. The analysis of the averaged ERPs focused on the previously indicated components of the average signal, with each component characterized by its amplitude, polarity (positive or negative), and latency. An ERP waveform consists of a series of peaks (here termed positive peaks) and troughs (negative peaks), but these voltage deflections reflect the sum of several relatively independent underlying, or latent, components. Isolating the latent components from the observable peaks and troughs of the waveform was challenging. The SubSearch game was designed to minimize latent components and to make sure that the evoked P300 was, as much as possible, a direct result of the experimental design. An important objective of the design was to separate the processes related to monetary gains and losses from possible confounding factors. The EEG does not only include ERPs but also other, “noisy” signals. The method we used to reduce the latter signals was signal averaging. All of the analyses featured epochs that were time-locked to the onset of the fixation signal that preceded the on-screen messages announcing reinforcers and punishers. Additionally, we examined the modulating effects of valence and magnitude on the ERP.
Figure 1: The timeline for the components of the ERP epoch.

Figure 1 depicts the sequence of events in the computer game. Each ERP epoch began with a blank screen that appeared simultaneously with scheduled delivery of a reinforcer or punisher. Five- hundred ms later, a fixation mark appeared on the screen. After another 500 ms had passed, it disappeared, and the gain or loss message appeared on the screen. It marked the onset of the P300 waveform. Analysis of the epoch began 500 ms previous to the fixation signal. Immediately following the presentation of the reinforcer (or punisher) message, which remained on the screen until the participant resumed the game, there was a 1000-ms delay until the tab at the bottom of the screen between the two cumulative counters began to blink. During this interval, the game was inoperative and remained so until the participant clicked the tab.
Behavioral data analysis
Thorndike [16] formulated the basic principle of the law of effect, which stated that actions followed by feelings of satisfaction are more likely to be repeated, but actions followed by feelings of annoyance are less likely Herrnstein [6,13] produced systematic work on behavioral choice involving schedules of reinforcement. He found that, over time, the proportion of responses to an alternative matched the proportion of reinforcers received for responding to that alternative. If twice as many reinforcers were provided to one of two alternatives, then, on average, twice as many responses were directed to that alternative once response allocation was stable. Herrnstein summarized this regularity as follows and termed it the matching law of distributed (ongoing) choice between alternatives:
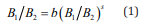
B refers to number of behavioral responses and R to the number of reinforcers that the responses produced. The two alternatives are designated by subscripts. This version of the matching law is actually a special case of the generalized matching law (GML; Baum, 1974):
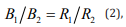
that is, b = s= 1.0.
The parameters in this power function reflect a bias (b) for one source of reinforcement over the other and sensitivity (s) to changes in the distribution of reinforcers between the two sources, respectively. Under logarithmic transformation, the GML becomes a linear equation:
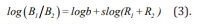
We used a procedure like that of Rasmussen and Newland and applied both the subtractive model as well as an indirect model to our results. The latter does not directly include punishers but, instead, represents the effect of punishers by variations in the effects of reinforcers. In other words, using this model, it is not necessary to include both reinforcers and punishers in the equation to measure their asymmetrical effect. All analyses were conducted using IBM SPSS Statistics 23 [13] and Microsoft Excel®. Measures included in the analyses were the number of responses (clicks) to the left and right alternatives (BL and BR) and the number of reinforcers provided by each (RL an d RR). The results were analyzed using equation 3. Asymmetry ratios were computed using antilogarithms of the bias estimates from the linear regressions for the conditions in which only reinforcers were used and those in which reinforcers and punishers were both used.
Results
Behavioral gain: loss asymmetry
Loss amplitudes were higher than gain amplitudes with an asymmetry ratio of 3.40. Table 2 contains the overall mean values of b for the No-punishers and With-punishers conditions, the 95% confidence intervals, and the asymmetry ratios for each session. Tables 3 and 4 contain the overall mean responses, obtained reinforcers, obtained punishers, and changeovers for the Nopunishers and With-Punishers conditions for the unpunished alternatives, respectively, for each of the participants. Table 5 is a summary of the overall estimated values of s and b in the Nopunishers and With-punishers conditions, and the asymmetry ratio for each participant.
Table 2: Overall Means and Standard Errors of the Values of b in No-Punishers and With-Punishers, 95% Confidence Intervals (CI), df, and Asymmetry Ratios Per Session.

Table 3: Means and Standard Errors of the Mean for Responses, Obtained Reinforcers, Obtained Punishers, and Changeovers (COs) for Each Participant in Conditions 1, 3, and 5 (No-punishers).

Table 4: Means and Standard Errors of the Mean for Responses, Obtained Reinforcers, Obtained Punishers, and Changeovers (COs) for Each Participant in Conditions 2, 4, and 6 (With- punishers).

Table 5: Overall Mean Estimated Values of s, b, and the Asymmetry Ratio in the No-punishers and With-Punishers Conditions for Each Participant.

Event related potentials
Figure 2: Grand Averaged VEP Waveforms During the Task. (A) Timeline of events in the SubSearch game protocol. (B) VEPs obtained at electrode F3 timed with the events in the SubSearch game. (C) VEPs obtained at electrode O1 timed with the events in the SubSearch game. Note the greater amplitude of the P300 ERP with losses compared to gains. Also shown is the mean reaction time (RT) for gains and losses. Approximate electrode placements appear in the head illustration at the right.

Figure 3: Overall Means and SEMs for Behavior and VEP Recordings. (A) Behavioral responding for gains and losses over the 7 sessions. (B-F) VEP components P100, N100, P200, N200, and P300 over the 7 sessions. The blue line represents the mean overall asymmetry ratio according to the right vertical scale.

Figure 2 displays the overall mean P300 waveform and other waveforms at electrode sites F3 (Panel B) and O1 (Panel C) for all participants according to the timeline. The approximate locations of the electrodes also appear. The reaction time (RT) was measured as the interval that began with the onset of the on-screen message and continued until the blinking tab was clicked. The mean RT was slightly shorter for the gain message (1512 ms) than for the loss message (1665 ms). In Figure 3, the overall mean amplitudes and standard errors are shown for each session: green bars for gains and red bars for losses. The blue lines represent the overall mean asymmetry ratio in each session. Five component waveforms appear, including the P300. The mean overall symmetry ratio for the P300 waveform was 1.99.
Correlation of the behavioral results with the electrophysiological results
Figure 4 displays the means of the asymmetry ratios derived from the two categories of data (behavioral on the x-axis and EEG on the y-axis) for each participant, together with the regression equation and R^2 (%VAC). Overall, the correlation coefficient was 0.852 and R^2 = 0.738.
Figure 4: Linear Regression of the Asymmetry Ratios for Each Participant Derived From the Behavioral Data (X-Axis) and From the EEG data (Y-Axis).

Discussion
Our results demonstrated a direct, robust correlation between behavioral and EEG measures of gain-loss asymmetry. The overall mean asymmetry ratio from our behavioral results was higher (M = 3.40) than that from the cognitive results previously cited [5,17] and from Rasmussen and Newland’s [14] behavioral study (M ≈ 3.0). For our EEG results, M = 1.99. One possible reason for the larger ratio from our behavioral results may be the use of the EMOTIV EPOC+® and the incumbency on participants to wear it throughout each session and, at the same time, to avoid unnecessary movements (which could have interrupted Bluetooth® interconnectivity or added noise to the EEG record). Notable aspects of the research method and data analysis included the use of the interdependent schedules of reinforcement, the inclusion of six different schedules within the same session, and the achievement of relative stability of participants’ performance within seven sessions [12,16], and that our participants were men, but there were 16 of them in our study and five in theirs. We, too, utilized a computer game as the behavioral task, though the on-screen presentation in our game was more complex than in theirs, as was the concatenation of six conditions within a session as opposed to their presentation of a single condition over consecutive sessions. Our use of interdependent VI VI schedules of reinforcement produced ratios of reinforcement consistently closer to the programmed ratios than in their study, where the disparity between programmed and actual ratios produced by conc VI VI schedules of reinforcement was substantial. Also, when a VI schedule of punishment was conjoined with the reinforcement schedule in their procedure, the scheduled rate of punishment was half that of the scheduled rate of reinforcement as opposed to being the same rate, as in our experiment. As with reinforcers, their results displayed a considerable disparity between the scheduled and actual rates of punishers.
Finally, our study included the measurement of EEG activity concurrent with participants’ distributed choice behavior. The temporal alignment of the EEG record with the record of behavior during the same SubSearch session was critical to demonstrating the correlation between the two sets of data. This requirement was fraught with unanticipated complications and required extensive revision and testing of the communication protocol between both computers (see the earlier description of the procedure) before it was sufficiently reliable for use in the research we reported. Perhaps the most noteworthy outcome of our study was the strength of the positive correlation between the behaviorally derived and EEGderived asymmetry ratios. Though no extant models account for that relationship, its demonstration invites further investigation. For the present, we have concluded that losses affect ongoing behavior in a distributed-choice procedure more intensely than gains do and that this differential effect may be quantified using the record of brain activity that occurs simultaneously with distributed choice behavior. It is as if the brain holds a mirror to externally observable behavior, or vice versa.
References
- Baum WM (1974) On two types of deviation from the matching law: Bias and undermatching. J Exp Anal Behav 22(1): 231-242.
- Bernat EM, Nelson LD, Steele VR, Gehring WJ, Patrick CJ (2011) Externalizing psychopathology and gain-loss feedback in a simulated gambling task: dissociable components of brain response revealed by time-frequency analysis. J Abnorm Psychol 120(2): 352-364.
- Chiappa KA (1997) Evoked potentials in clinical medicine (3rd), New York, NY: Lippincott Williams & Wilkins, US.
- Ferster CB, Skinner BF (1957) Schedules of reinforcement. New York NY: Appleton- Century-Crofts, USA.
- Gehring WJ, Willoughby AR (2002) The medial frontal cortex and the rapid processing of monetary gains and losses. Science 295(5563): 2279-2282.
- Herrnstein RJ (1961) Relative and absolute strength of response as a function of frequency of reinforcement. J Exp Anal Behav 4: 267-272.
- Kahneman D (2011) Thinking, fast and slow. New York, NY: Farrar, Straus and Giroux, USA.
- Kahneman D, Knetsch J, Thaler R (1991) Experimental tests of the Endowment Effect and Coase Theorem. Journal of Political Economy 98(6): 1325-1348.
- Kahneman D, Tversky A (1984) Choices, values, and frames. American Psychologist 39: 341-350.
- Kahneman D, Tversky A (1979) Prospect theory: An analysis of decision under risk. Econometrica 47: 263-291.
- Luck SJ (2005) An introduction to the event-related potential technique. Cambridge, MIT Press, MA, USA.
- Polich J (2007) Updating P300: An integrative theory of P3a and P3b. Clinical Neurophysiology 118(10): 2128-2148.
- Rachlin H, Laibson DI (1997) The matching law: Papers in psychology and economics. Cambridge, Harvard University Press, MA, USA.
- Rasmussen EB, Newland MC (2008) Asymmetry of reinforcement and punishment in human choice. J Exp Anal Behav 89(2): 157-167.
- Sokol Hessner P, Rutledge RB (2019) The psychological and neural basis of loss aversion. Current Directions in Psychological Science 28(1): 20-27.
- Thorndike EL (1911) Animal intelligence. Macmillan, New York, NY, USA.
- Yeung N, Sanfey AG (2004) Independent coding of reward magnitude and valence in the human brain. Journal of Neuroscience 24(28): 6256-6264.


-

Mark E Smith
Bio chemistry
University of Texas Medical Branch, USA -

Lawrence A Presley
Department of Criminal Justice
Liberty University, USA -

Thomas W Miller
Department of Psychiatry
University of Kentucky, USA -

Gjumrakch Aliev
Department of Medicine
Gally International Biomedical Research & Consulting LLC, USA -

Christopher Bryant
Department of Urbanisation and Agricultural
Montreal university, USA -

Robert William Frare
Oral & Maxillofacial Pathology
New York University, USA -

Rudolph Modesto Navari
Gastroenterology and Hepatology
University of Alabama, UK -

Andrew Hague
Department of Medicine
Universities of Bradford, UK -

George Gregory Buttigieg
Maltese College of Obstetrics and Gynaecology, Europe -

Chen-Hsiung Yeh
Oncology
Circulogene Theranostics, England -
.png)
Emilio Bucio-Carrillo
Radiation Chemistry
National University of Mexico, USA -
.jpg)
Casey J Grenier
Analytical Chemistry
Wentworth Institute of Technology, USA -

Hany Atalah
Minimally Invasive Surgery
Mercer University school of Medicine, USA -

Abu-Hussein Muhamad
Pediatric Dentistry
University of Athens , Greece
